广告
非洲猪瘟(ASF)是由非洲猪瘟病毒(ASFV)引起的猪的一种高度接触性传染病,对家猪致死率可达100%,严重威胁我国及世界养猪业可持续发展,目前尚无确实安全有效的预防用疫苗。本文总结了ASFV的流行历史与态势、生物学特性与复制、感染与致病机制、与靶细胞的相互作用以及适应性免疫应答,并讨论了相关预防与控制措施,以期为我国ASFV研究和防控提供帮助。
非洲猪瘟(African swine fever,ASF)是威胁全球养猪业的“头号杀手”,为世界动物卫生组织法定报告动物疫病,在我国为I类动物疫病。该病是由非洲猪瘟病毒(African swine fever virus,ASFV)引起的 一种高度接触性传染病。各种年龄家猪均易感,最急性型、急性型和亚急性型感染对家猪具有高度致死性,患猪病死率可达100%。
1 ASFV流行历史与态势
ASFV在世界范围内的流行已逾百年。1921年, 病毒学家 Montgomery 首次报道了肯尼亚ASF疫情,自此该病在撒哈拉沙漠以南的非洲国家广泛流行。20世纪50年代后期,基因 I 型ASFV传入欧洲、南美和加勒比海周边的国家。得力于严格的生物安全控制和扑杀措施,很多国家和地区于20世纪90年代中期有效控制或根除了ASF。2007年左右,基因II型ASFV传入高加索地区的格鲁吉亚、亚美尼亚和俄罗斯,此后10年间,ASFV扩散至欧洲东部和中部、俄罗斯中部及远东地区。
自2018年8月,我国暴发首例由基因II型ASFV毒株引起的ASF疫情 ,并迅速扩散至除台湾之外的所有省、市、自治区,重创生猪产业。疫情还波及周边国家和地区,包括韩国、越南、蒙古、柬埔寨、朝鲜、老挝和缅甸等,整个东亚地区ASF风险持续攀升。自2018年传入我国后,ASF在我国的流行大致经历了4个阶段。
1)2018—2019年以基因II型强毒株的流行为主,感染猪致死率可达100%。
2)此后基因缺失毒株在田间出现,遗传多样性逐步增加,该类毒株毒力减弱,可引起慢性感染或无症状感染,感染性强,排毒不规律。
3)至2021年左右,基因I型毒株在田间出现,并与基因II型毒株发生混合感染,
4)其后出现I/ II型重组毒株,进一步加剧了我国 ASFV 毒株的多样性和复杂性。
更为重要的是,利用基因II型毒株制备的减毒疫苗不能对重组毒株提供交叉保护。目前,国内ASF流行毒株呈现“多谱系、多时空” 特点,给ASFV防控技术研发带来了新的挑战。
2 ASFV生物学特性与复制
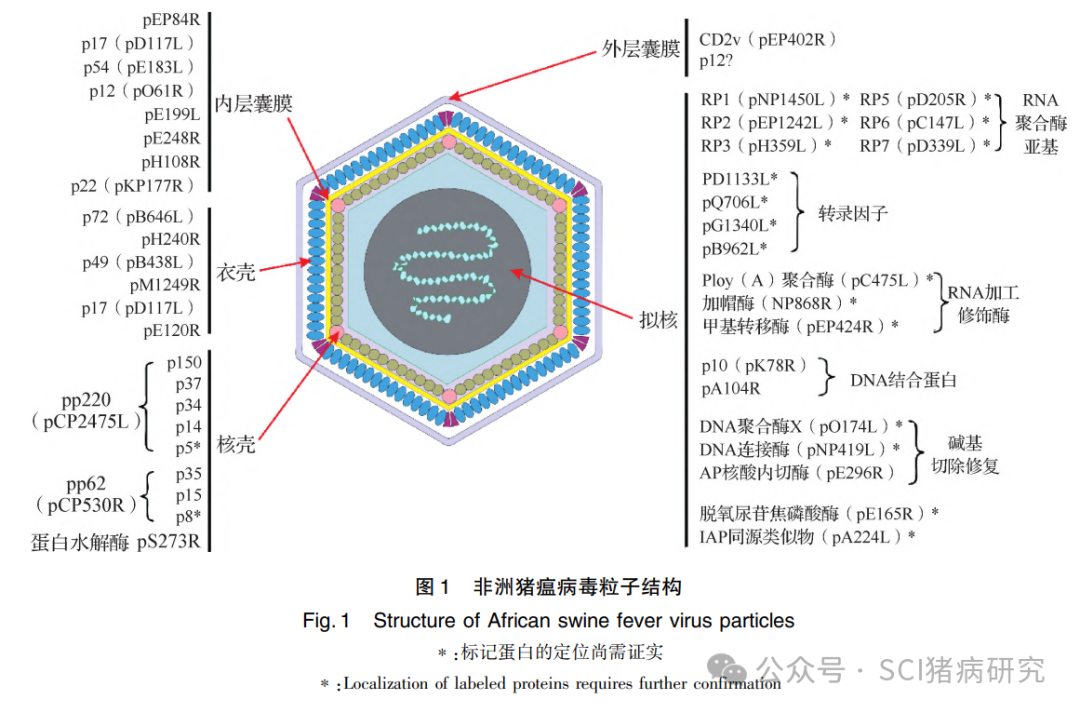
ASFV为一类有囊膜、大型、线性双链DNA病毒,归属非洲猪瘟病毒科非洲猪瘟病毒属,且为该科唯一成员,也是目前发现的唯一能在节肢动物中复制并通过节肢动物传播的虫媒DNA病毒。成熟的ASFV粒子呈二十面体形态,直径约250nm,具有5层结构,由内及外分别为拟核结构、核壳、内层囊膜、衣壳和外层囊膜[图1]。形态结构上,ASFV为经典病毒和巨病毒(Giant viruses,GV)之间的过渡病毒,既具有经典病毒的外层囊膜结构,又享有GV所独有的内膜结构。ASFV成熟的病毒粒子和缺少最外层囊膜的病毒粒子均具有感染性。ASFV 多数毒株具有红细胞吸附特性( Hemadsorption,HAD),有利于病毒的扩散和传播。ASFV基因组庞大,全长在160~200 kb,其长度差异主要源于多基因家族(Multigene families,MGFs) 基因数目的变化。ASFV编码超过150种蛋白质,从功能上可分为四大类,包括病毒结构蛋白和参与病毒粒子形成的蛋白(至少有54种)、参与病毒基因组复制和转录的酶类及辅助因子、参与宿主免疫和代谢调控的蛋白,以及由MGFs编码的病毒蛋白 。目前,大部分病毒蛋白功能仍未知,庞大的编码数量给保护性抗原基因和病毒毒力基因的筛选带来巨大挑战。
2.1 毒株毒力分型与持续性感染
ASF自然感染潜伏期为4~19d,实验条件下为2~5d,潜伏期长短与病毒遗传背景、接种剂量和感染途径有关。临床症状取决于毒株毒力、感染剂量和感染途径。
1)高毒力和中等毒力毒株引起的临床症状以猪只发热和网状内皮组织广泛性出血为主。高毒力毒株主要引起最急性或急性型 ASF,致死率可达90%~100%;
2)中等毒力毒株主要引起亚急性感染,致死率在成年动物为20%~40%,幼年动物为70%~80%;
3)低毒力毒株对家猪通常不致死,可引起慢性或无症状感染,感染猪只出现繁殖障碍和生长性能下降,部分表现为皮肤坏死和溃疡、关节炎以及呼吸道症状等。
耐过猪可长时间带毒,ASFV可与抗体同时存在于感染猪血液中,持续6个月至数年。低毒力毒株能够在猪体内建立持续性感染,感染猪只持续或间歇性向外排毒,成为重要传染源,是ASF根除计划中的关键对象。
2.2 遗传变异
ASFV 具有较高变异性,变异主要集中于基因组左右两侧的可变区,左边可变区(Left variable regions,LVR) 约35kb,右边可变区(Right variable region, RVR) 约15kb,其主要含有5个MGFs。MGFs 基因区域常发生多重拷贝基因的缺失或插入, 与病毒毒力和宿主范围 (蜱) 等有关;该区域也是细胞适应毒株基因缺失发生的部位,缺失长度可高达24.5kb,能增加毒株对细胞的适应性。ASFV基因组中部为保守区域(Central conserved region, C区), 长度在125kb左右。ASFV基因型和亚型的划分基于基因组中部保守区域中心可变区(Central variable region,CVR)的微小变异。基于B646L基因的遗传演化分析,ASFV存在至少24种基因型。欧洲主要流行基因I型和II型,亚洲流行毒株多为基因II 型,所有已知的24种基因型都能在撒哈拉以南的非洲地区检测到。根据ASFV CD2v血凝特性,可将ASFV分为至少8个血清群,型间或群间免疫交叉保护能力相对有限。
我国初期流行的ASFV毒株与2017年在俄罗斯流行的毒株(KY963545)以及 2007 年在格鲁吉亚流行的毒株高度同源,均为基因 II 型。随后, 基于全基因组监测分析,陆续发现了一些“人工缺失株”和自然基因缺失毒株的流行,缺失区域涵盖23个基因, 长度可达10 000bp, 主要集中于 MGF100、 MGF110、MGF360家族和 EP153R / EP402R 基因,这些毒株是我国 ASF 防控中面临的新污染源和传染源。与强毒株相比,基因缺失毒株毒力减弱、隐敝性强,感染猪排毒不规律,影响养殖场对ASFV有效的早期监测和检测。近期, 田间出现了I型毒株,以及I 型与II型的重组毒株。其中,重组毒株能突破基因II型减毒疫苗诱导的免疫保护屏障。此外,一些关键毒力基因的高频缺失以及毒株的重组可导致ASFV致病性、潜伏期和免疫逃避能力等多因素改变。持续性感染促进变异和重组发生,导致毒株多样化进一步加剧。
2.3 病原生态学特性
野猪和家猪为ASFV的自然宿主,猪只之间的主要传播方式为皮肤和体液的接触传播。非洲野猪(如疣猪、大森林猪和灌木猪等)对ASF具有极强的抵抗力,一般不表现临床症状,而家猪和欧洲野猪感染,表现从急性到慢性的各种临床症状。ASFV自然传播链复杂,涉及野猪、蜱类和家猪。目前发现的所有软蜱类均对ASFV易感,且ASFV能在软蜱体内存在并保持对猪的感染性长达数年,因此,存在软蜱的地区,一旦感染ASFV就很难彻底净化。
2.4 ASFV复制周期
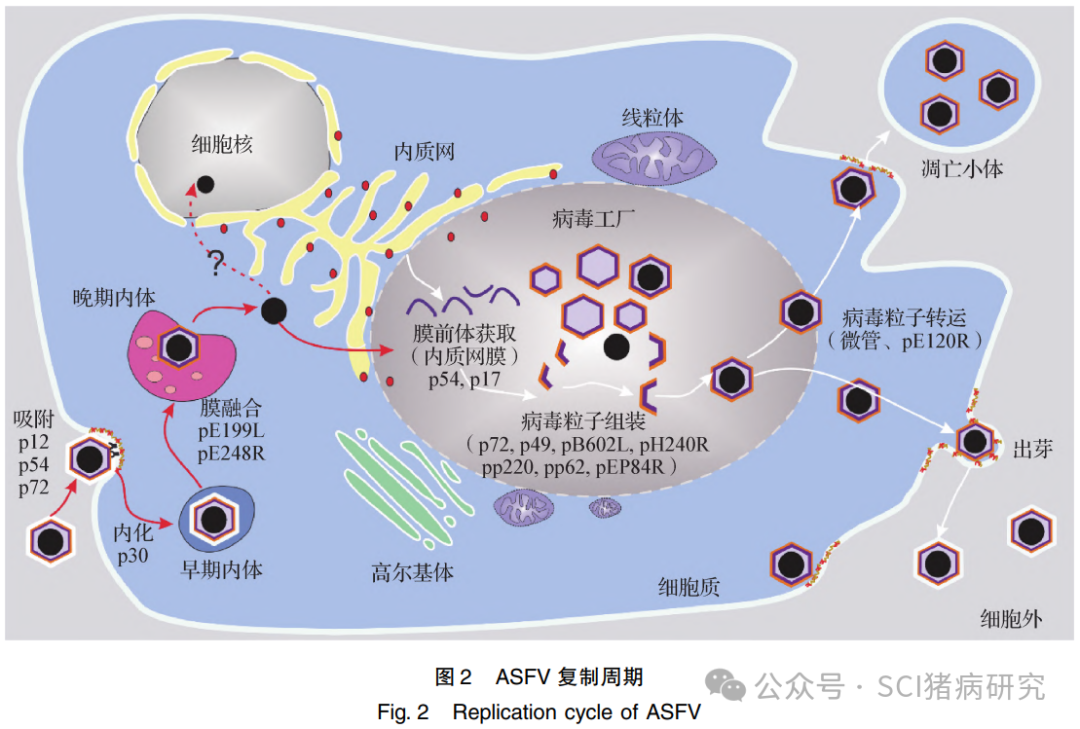
ASFV 主要靶向单核巨噬细胞系统,目前已经鉴定的参与ASFV入侵过程的相关宿主分子有富含半胱氨酸的B型清道夫受体 ( Scavenger receptor cysteine-rich type 1 protein M130,CD163)、白细胞共同抗原(Leukocyte common antigen,LCA)、主要组织相容性复合体 I 型(Major histocompatibility complex class I,MHC-I)、CD1d 分子、AXL氨酸蛋白激酶受体(AXL receptor tyrosine kinase,AXL) 和磷脂酰丝氨酸( Phosphatidylserine, PS)等,但是关键入侵受体尚未完全解析。研究还发现, ASFV结构蛋白p12(pO61R)、p72 (pB646L)、p54 ( pE183L) 和 p30 ( pCP204L) 等参与病毒吸附和内化过程。ASFV通过多种途径入侵宿主细胞,包括肌动蛋白驱动的巨胞饮、网格蛋白介导的内吞途径和凋亡小体介导的胞葬途径等。其中,凋亡小体介导的胞葬途径可以逃逸抗体的中和作用,可能是 ASFV 在猪体内最有效的传播和入侵策略,但不管哪种入侵途径,ASFV 最终到达内体系统完成脱衣壳, 随后病毒内膜蛋白 (pE248R 和 pE199L 等)介导病毒内膜与细胞内体发生膜融合,释放包裹病毒基因组的“裸”核心至细胞质。ASFV主要在细胞质建立病毒工厂, 集病毒转录、复制、蛋白质合成和病毒粒子装配于一体。有研究表明,ASFV 基因组复制可能存在短暂核内期,但需要更多证据证实。组装好的细胞内病毒粒子被转运至细胞膜,以出芽或诱导形成凋亡小体的方式释放(图2)。
2.5 免疫逃逸与抑制特性
ASFV感染能够潜在影响单核巨噬细胞的吞噬杀伤、抗原递呈和免疫调节等诸多功能,进而影响宿主天然免疫和适应性免疫应答。同时,ASFV 编码大量病毒蛋白用于拮抗 I 型干扰素信号通路,调控细胞凋亡,抑制宿主免疫相关基因转录和翻译,以及调控炎症因子表达等。ASFV编码几乎整套用于自身基因转录和复制的蛋白,且病毒复制和组装主要发生在细胞质内的病毒工厂,对细胞周期的依赖性较小。
3 ASFV感染与致病机制
经口感染时,ASFV首先在扁桃体和下颌淋巴结的单核细胞和巨噬细胞中复制,随后扩散至二级复制场所,包括淋巴结、骨髓、脾脏、肺脏、肝脏和肾脏。病毒血症通常出现在感染ASFV后4~8d, 可持续数周或数月或更久。II型ASFV强毒株感染的临床病理学特征主要表现为以下3点:(1) 高热(40.5~42.0℃)和广泛性出血。感染猪只的心脏、肺脏和肾脏等实质脏器有严重出血症状,淋巴结肿大并呈严重弥漫性出血。微血管的损伤和出血是特征性病理变化,出血最严重的组织主要是淋巴相关组织。这种出血机制尚不十分清楚, 但应与多种因素有关,包括感染过程中局部炎症因子的上调、循环中血小板数量的减少和病毒感染诱导的弥散性血管内凝血等。在感染后期,血管内皮细胞中能检测到病毒抗原的存在。但也有学者认为,病毒感染诱导的内皮细胞损伤可能不占主要地位,因为大多数血管病变发生在病毒感染内皮细胞以前。此外,不排除抗原-抗体复合物介导的变态反应对此过程的影响。(2)淋巴相关组织的破坏和淋巴细胞的大量减少。这种损伤主要发生在T淋巴细胞分布区域,但还没有证据表明病毒能够在淋巴细胞内复制。淋巴细胞减少可能与病毒感染诱导的非感染细胞的凋亡有关。感染的巨噬细胞分泌的炎症因子,如肿瘤坏死因子-α (Tumor necrosis factor-α,TNF-α)可能是引起淋巴细胞凋亡的重要机制。目前支持这一假说的证据有2个:主要是感染ASFV的巨噬细胞周围的淋巴细胞发生凋亡;用病毒感染巨噬细胞的培养上清(不含病毒粒子)处理淋巴细胞,可诱导淋巴细胞凋亡。此外,感染ASFV猪只的淋巴细胞对有丝分裂原凝集素刺激没有反应,表明ASFV感染不仅导致淋巴细胞数量减少,而且对其功能产生了影响。(3)充血性脾肿大。常见于ASF急性或亚急性感染,主要发生在高毒力毒株感染的猪群。脾脏肿胀至正常大小的3~6倍,颜色变暗,质地变脆。有研究认为,红髓脾索上巨噬细胞的感染、凋亡或坏死使脾索血管的基底膜暴露,激活了凝血系统,并导致纤维素沉积,从而导致红细胞在脾索上沉积。同时, 由于巨噬细胞大量死亡,脾脏血液的过滤和清除功能受损,大量血液淤积于脾脏。组织缺氧也间接促进了脾脏组织淋巴细胞的凋亡或坏死。
与基因 II 型强毒株相比,ASFV基因I型毒株的毒力明显降低,因此感染 I型ASFV的猪只主要为慢性感染,感染猪只血液中的病毒载量低,部分猪只甚至在观察期内未能检出,猪体排毒不规律。基因 I 型ASFV毒株感染的猪只较少发生炎症因子风暴,表现为白细胞介素10( Interleukin-10,IL-10)、C-X-C基序趋化因子配体8(C-X-Cmotif chemokine ligand 8,CXCL8)等细胞因子下调,促炎因子C-X-C基序趋化因子配体10(C-X-C motif chemokine ligand 10,CXCL10)上,γ-干扰素(Interferon-γ,IFN-γ)无变化,白细胞介素6(Interleukin-6,IL-6)和α-干扰素 (Interferon-α,IFN-α)上调。
4 ASFV与靶细胞相互作用
单核-巨噬细胞是宿主天然免疫的重要组成部分,具有至少5大功能,包括吞噬和杀伤功能、抗原递呈功能、细胞因子和炎症因子分泌与调节功能、可结晶片段(Fragment crystallizable,Fc)和补体受体介导的免疫杀伤与调理功能,以及组织修复功能 (分泌促生长因子)。因此,阐明 ASFV感染对靶细胞(单核-巨噬细胞)的影响是解析病毒致病机理的重要环节。围绕 ASFV与猪单核-巨噬细胞相互作用展开了系列研究,主要进展如下。
4.1 调控巨噬细胞凋亡4.2 调控宿主细胞的转录和翻译
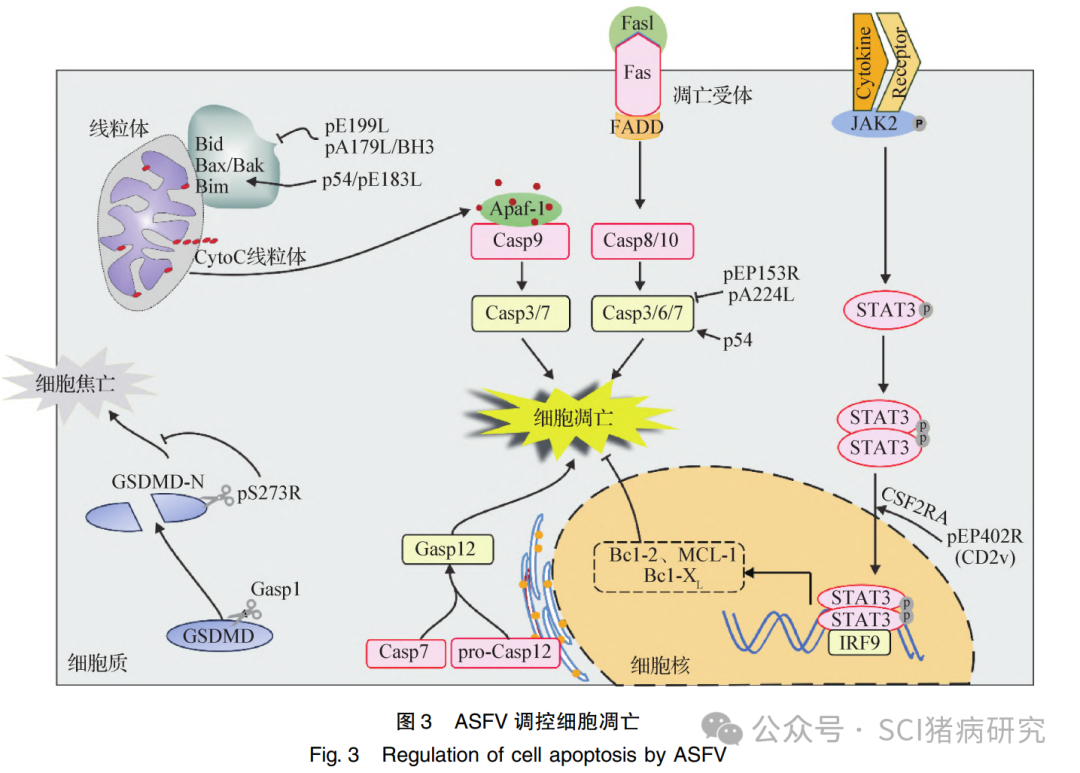
ASFV 在复制早期抑制细胞凋亡,促进细胞存活,利于病毒复制;在感染后期诱导细胞凋亡,促进子代病毒释放。目前,已鉴定出至少4种病毒抗凋亡蛋白,包括 pA179L、 pA224L、pEP153R 和 pEP402R (图3)。其中,pA179L 是宿主B淋巴细胞瘤-2(B-cell lymphoma-2,Bcl-2)蛋白家族类似物;pA224L 为宿主抗细胞凋亡凋亡抑制蛋白(Inhibitor of apoptosis proteins, IAPs)的同源类似物;pEP153R 是一种C型凝集素, 通过抑制 Caspase 3的活性抑制凋亡;pEP402R(即囊膜糖蛋白 CD2v)通过与宿主蛋白集落刺激因子2受体α亚基 ( Colony stimulating factor 2 receptor fubunit alpha,CSF2RA)相互作用激活 Janus 激酶2-信号传导和转录激活蛋白3 ( Janus kinase 2-signal transducerand activator of transcription 3, JAK2- STAT3)信号通路,抑制宿主细胞凋亡。缺失编码上述抗凋亡蛋白的基因并不影响病毒毒力及其诱导的细胞凋亡,表明病毒还编码其他抗细胞凋亡蛋白。此外,病毒蛋白酶pS273R可以非经典切割消皮素D( Gasdermin D,GSDMD),抑制细胞焦亡。在感染后期,ASFV能够激活内质网通路,内质网内 Caspase 12 被显著活化,进而诱导细胞凋亡。还有研究表明,ASFV 结构蛋白 p54 具有诱导细胞凋亡活性,pA179L抑制细胞凋亡的同时诱导细胞坏死,利于新生病毒粒子的释放,囊膜蛋白pE199L 可以诱导线粒体依赖性细胞凋亡 (图3) 。目前,ASFV是否编码其他诱导细胞凋亡的病毒因子还有待探究。
4.2 调控宿主细胞的转录和翻译
ASFV在复制过程中利用宿主细胞转录和翻译相关物质资源,确保病毒基因的有效转录和翻译,同时抑制宿主细胞基因的转录和翻译,进而抑制宿主细胞抗病毒反应。目前,已鉴定出5种影响宿主基因转录和翻译的重要病毒蛋白, 包括pA238L、pMGF110-7L、pE66L、pK205R 和 pI73R 。pA238L蛋白为宿主核因子kappa-B激酶抑制剂(Inhibitor of nuclear factor kappa-B kinase,IκB)的同源类似物,通过抑制核因子 κB(Nuclear factor kappa-B,NF-κB)激活,抑制多种基因转录,尤其是编码参与炎症反应和免疫信号转导的细胞因子和趋化因子;pA238L 蛋白还可与钙神经素结合,抑制依赖活化的T细胞核内因子(Nuclear factor of activated T cells,NFAT)基因,如编码白细胞介素2(Interleukin-2,IL-2)、白细胞介素 4(Interleukin-4,IL-4)、粒细胞-巨噬细胞集落刺激因子(Granulocyte-macrophage colony stimulating factor, GM-CSF) 等基因的转录。ASFV编码蛋白pE66L、pK205R和 pMGF110-7L 等可通过激活蛋白激酶 R(Protein kinase R,PKR) 和蛋白激酶 R 样内质网激酶( Protein kinase R-like endoplasmic reticulum kinase, PERK) 通路抑制细胞翻译。pI73R可选择性抑制宿主细胞 mRNA 的核输出。
ASFV对宿主细胞翻译机器的调控体现在以下3个方面:(1)劫持宿主翻译机器。ASFV可招募翻译相关因子,如真核起始因子 2α(Eukaryotic initiation factor-2α,eIF2α)、真核起始因子 3B(Eukaryotic initiation factor-3B,eIF3B)、真核起始因子 4E (Eukaryotic initiation factor-4E,eIF4E)、真核起始因子 4F(Eukaryotic initiation factor-4F,eIF4F)、真核起始因子4G(Eukaryotic initiation factor-4G,eIF4G)、真核 翻译延伸因子 2(Eukaryotic translation elongation factor 2,eEF2)和核糖体 P 蛋白(Ribosomal P protein) 等至病毒复制工厂,促进自身翻译;(2) 编码病毒蛋白, 促进宿主mRNA的降解 (ASFV脱帽酶),或者抑制宿主细胞mRNA核输出,进而抑制宿主蛋白合成(pI73R);(3) 调控翻译起始因子eIF2α的磷酸化。pDP71L蛋白通过招募去磷酸化酶生长停滞和 DNA 损伤诱导基因 34(Growth arrest and DNA damage-inducible gene 34,GADD34)对 eIF2α-P 进行去磷酸化,但 pDP71L可能不是该过程唯一的病毒调控因子,因为缺失 pDP71L编码基因的病毒仍然能够阻止eIF2α 的磷酸化。

4.3 调控炎症因子的表达和分泌
过度激活的单核细胞或巨噬细胞分泌的炎症因子可能是病毒诱导病理损伤的关键环节。已有研究表明,在急性ASFV感染猪只中,血清和组织中的炎症介质 TNF-α、IL-1α 和 IL-1β 呈升高趋势,且与猪只的高热呈正相关。还有研究表明,炎症因子的浓度和升高水平并不是均一的,而是与 ASFV 在体内传播过程中到达的器官密切相关。局部高浓度炎症因子的分泌可能参与病毒感染诱导的病理损伤。有研究发现,发生凋亡的淋巴细胞主要存在于 ASFV 感染的巨噬细胞周围,因而认为,ASFV 感染细胞分泌的炎症因子可能是诱导非感染淋巴细胞凋亡的重要因素。此外,高浓度的炎症因子还可引起局部血管通透性增强,长期高浓度的炎症因子还可引起器官衰竭。目前,对ASFV如何调控炎症因子表达的分子机制研究较少。研究鉴定出多个调控炎症因子的病毒蛋白,其中抑炎因子包括 pS183L、pE199L、pO61R、pH240R 和pI7L,促炎因子包括pI226L、pA151R、pNP419L、 pQP383R、pMGF505-7R、pMGF300-2R 和pMGF300-4L。pH240R可以靶向增强核苷酸结合寡聚结构域样受体蛋白 3(Nucleotide-binding oligomerization domain-like receptor protein 3,NLRP3)炎症小体介导的炎症应答,进而增强病毒致病力;敲除 ASFV 的MGF300-4L、MGF300-2R 和 MGF505-7R 基因可以显著增强炎症因子(IL-1β和TNF-α) 的表达。此外,研究还证实,ASFV 感染可下调宿主抗炎症因子补体C1q肿瘤坏死因子相关蛋白3 (C1q and tumor necrosis factor related protein 3, C1QTNF3),进而激活炎症反应,具体机制有待进一步探究。
4.4 调控干扰素信号通路
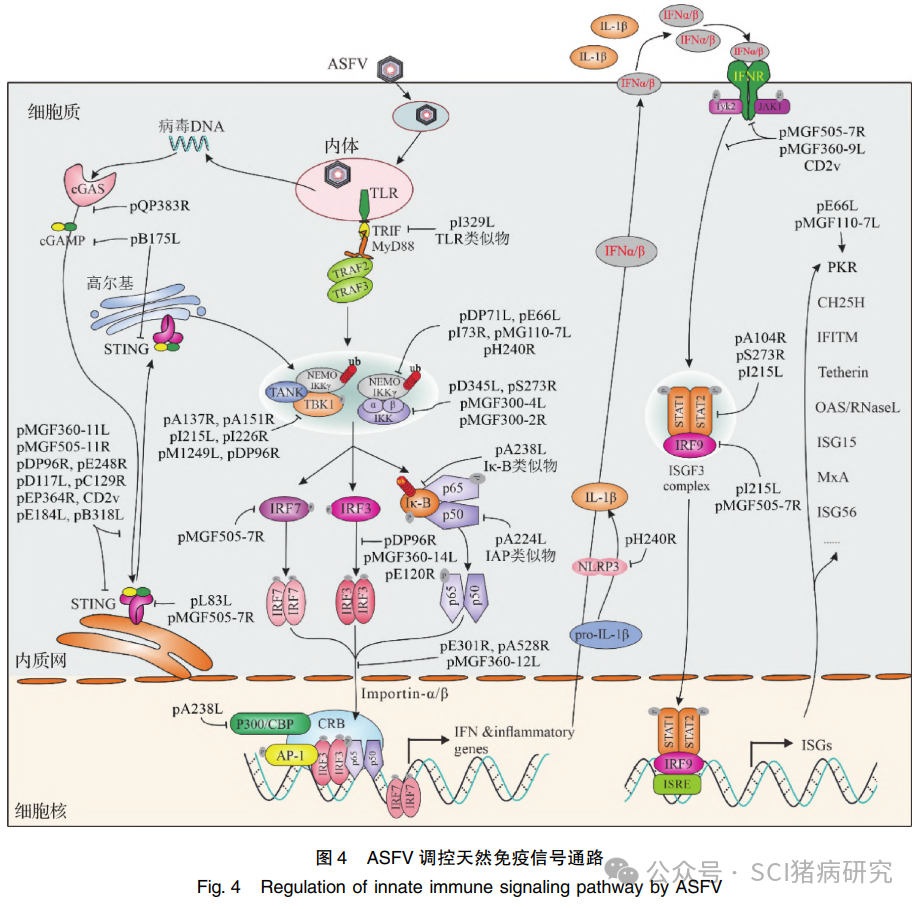
I型干扰素信号通路是宿主抗病毒感染的重要防御机制。ASFV基因组两侧高变区域编码的MGF蛋白(如pMGF100、110、300、360和530等)除了与病毒组织细胞嗜性相关之外,大多具有拮抗宿主细胞干扰素信号通路的功能,如pA276R(pMGF360-15R)、pA528R、pMGF110-7L、pMGF360-9L、pMGF360-11L、 pMGF360-12L、pMGF360-14L、pMGF300-2R、pMGF300-4L、pMGF505-7R和pMGF505-11R等。此外,研究还鉴定了其他具有抑制干扰素信号通路的重要病毒蛋白, 如 pA104R、pA137R、pA151R、pA224L、pA238L、pB175L、pB318L、pC129R、pD117L、pD345L、pDP71L、pDP96R、pE120R、pE184L、pE248R、pEP364R、pEP402R、pL83L、pH240R、pQP383R、pS273R、pI226R 和 pI215L 等。上述分子主要作用于 cGAS-STING 和下游 JAK-STAT 信号通路等(图4),调控关键信号转导蛋白的丰度、修饰和亚细胞定位,进而抑制 I 型干扰素信号转导。目前,通过缺失上述拮抗干扰素反应的关键病毒因子,获得了多株毒力致弱、且可为亲本毒株提供良好免疫保护作用的疫苗候选毒株, 但尚需大量试验分析其安全性。
4.5 对巨噬细胞吞噬杀伤和抗原递呈功能的影响
有关 ASFV感染对巨噬细胞吞噬和杀伤功能影响的研究甚少,尤其是ASFV感染对巨噬细胞吞噬杀伤相关模式识别受体、信号转导分子、吞噬杀伤效应分子和氧化代谢酶类等的影响和调控机制。有关 ASFV 感染是否能下调巨噬细胞表面MHC-I 或II类分子表达仍有争议,可能存在毒株特异性。根据功能和活化状态,可将巨噬细胞分为 M1和M2两种亚型。M1型主要功能为吞噬、加工及递呈抗原和杀伤作用,发挥正向免疫调控作用;M2型的抗原递呈和杀伤能力较弱,主要通过分泌IL-10 和TGF-β 等抑制性细胞因子表达,抑制淋巴细胞增殖,发挥负向免疫调节作用。有研究表明,巨噬细胞M1型极化可导致 ASFV复制能力下降,但M2型极化并不影响ASFV的易感性。精氨酸 1 (Arginase-1, Arg-1)作为M2型巨噬细胞的重要标志分子,在 ASFV感染的巨噬细胞中表达显著上调, 但ASFV感染调控巨噬细胞极化的具体机制尚不清楚。
4.6 与细胞信号通路相互作用调控病毒复制
相关研究表明,ASFV可激活内质网应激-激活转录因子6(Activating transcription factor 6,ATF6)和 PERK 信号通路, 并鉴定了2 个病毒蛋白 pK205R 和 pMGF110-7L 参与这一过程。有关 ASFV 感染对细胞自噬流的影响、调控机制和生物学意义尚不清楚, 仅体外研究显示, ASFV蛋白 pA179L和pE199L分别与Bcl-2互作蛋白(Bcl-2 interacting protein,Beclin)和吡咯啉-5-羧酸还原酶 2(Pyrroline5-carboxylate reductase 2,PYCR2) 相互作用调控细胞自噬。此外,ASFV 可以通过抑制 eIF2α 磷酸化和病毒编码的半胱氨酸蛋白酶 pS273R 裂解 G3BP 应激颗粒组装因子1(G3BP stress granule assembly factor 1,G3BP1)2 种策略抑制抗病毒应激颗粒的形成,进而促进病毒复制。还有研究发现,ASFV 感染可以利用蛋白质泛素化和泛素-蛋白酶体系统促进自身复制。
5 适应性免疫应答
感染ASFV的幸存猪只产生可抵抗同源毒株攻击的保护性免疫,但是对异源毒株的攻击不能提供有效交叉保护。目前,对同源免疫的保护机制知之甚少,但细胞免疫似乎占主导地位。利用感染猪只或其他动物的抗血清进行被动免疫能够提供部分保护,能延迟感染猪只的发病时间,缓解临床症状, 降低病毒血症水平, 提高猪只的存活率;然而,耗竭 CD8+T 细胞可极大降低弱毒株诱导的同源免疫保护效率。近期研究发现, ASFV 可利用凋亡小体进行入侵和胞间传播,以逃逸抗体中和作用,该研究部分阐释了ASFV“中和抗体低下/无效”现象。利用单细胞测序对基因II型ASFV强毒株感染猪只的分析结果显示,ASFV感染可导致自然杀伤(Natural killer,NK)细胞、巨噬细胞和T细胞数量急剧减少,相反,单核细胞数量大幅度增加;进一步研究发现,ASFV主要感染脾脏单核细胞和巨噬细胞,且感染的单核细胞为非经典的未成熟的CD14-单核细胞,该类细胞中干扰素刺激基因(Interferon-stimulated genes,ISGs) 表达水平和细胞凋亡水平均较低。
6 ASFV预防与控制
目前,尚无针对ASF的药物和安全有效的预防用疫苗。ASFV灭活苗不能诱导保护性免疫;减毒活疫苗可对同源毒株提供较好保护作用,但安全性存在不足。强化猪场生物安全仍是目前ASF防控的关键。应建立合理的猪场生物安全防控体系和风险评估系统,严控“猪、人、料、运输工具、物资、环境” 6个关键环节,严把引种关,切忌引进 ASFV 带毒猪;加强监测和检测,通过分子诊断和血清学监测精准清除和严格处置ASF疫情, 防止ASFV 在场内扩散和传播,同时对发病猪、感染猪、耐过猪及其污染物进行严格的无害化处理,禁止进入流通环节;控制 ASFV自然储藏宿主,如软蜱和野猪等,防止其与家猪接触;加强 ASFV跨境传入风险管理,加强控制生猪和猪副产品流通,进行产品检疫监测,对国际机场和港口中的垃圾和食物残余等进行焚烧处理。总体上,ASFV防控与净化策略因情形而异,与所在地区的政治、经济状况、文化、气候和流行病学情形等密切相关。有条件的生猪养殖地区和养殖企业应加大力度建设ASF无疫区,并加强区域协同,为ASF的净化和根除奠定基础。
7 小结与展望
ASF是当前威胁我国及世界养猪业可持续发展的重大疫病,极大威胁了生猪供应链安全和社会经济稳定。ASFV基因组庞大,编码蛋白众多,病毒粒子结构复杂,人们对其认知仍极为有限。当前,制约ASFV防控和安全高效疫苗研制背后的若干基础科学问题仍未得到解决,包括ASFV功能基因组学与编码蛋白功能、入侵与复制机制、感染与病理损伤机制、免疫抑制与逃逸机制、免疫保护机制等。此外,近期基因缺失变异毒株和I/II型重组毒株的出现给我国ASF疫情防控带来严峻挑战,亟待进一步加强其病原生态学和流行病学研究,揭示毒株遗传演化的时空规律、传播特性和致病特性。加强ASFV病原生物学与免疫致病机制研究可为防控策略制定、安全高效疫苗设计和抗病毒药物研发提供思路和依据,进而更有效地保障养殖安全、生态安全和食品安全。