
广告
猪繁殖与呼吸综合征病毒(PRRSV),俗称猪蓝耳病毒,对中国养猪业造成持续的影响。关于PRRSV的分类,根据抗原性,PRRSV分为PRRSV-1(欧洲型)和PRRSV-2(美洲型),但两种基因型的核苷酸序列同源性仅为50-70%。根据ORF5核苷酸序列的遗传演化分析和PRRSV的全球分类系统,将PRRSV-1分为3-4个亚型。PRRSV-2可分为9个亚型:从谱系1到谱系9。虽然PRRSV-2在中国的流行占主导地位,但PRRSV-1毒株在中国也已经存在了20多年。
近年来,PRRSV-1在中国的流行受关注较少。但不可否认,PRRSV-1已蔓延到中国的23个省、市和自治区。来自哈尔滨兽医研究所田志军和张洪亮研究团队将中国PRRSV-1毒株在中国的流行情况、基因组特征与演化、致病性、疫苗研发以及预防与控制措施做了总结性分享。

迄今为止,很少有研究调查呼吸道疾病与猪肉品质之间的关系。研究发现,与没有病变的胴体相比,肺部病变严重的胴体发生肉质异常(倾向于DFD 和 PSE肉)的风险明显更高。此外,强有力的证据表明:肺部病变与肉类感官颜色评分的变化、pH依赖性肉类参数(如:系水力、滴水、解冻和烹饪损失)之间存在关联。
在屠宰场,主要采用传统手工方法对肺部大体病变和胸膜肺炎进行分级,有公司可提供数字化评估软件。软件通常集成了传统方法,如:Madec和Kobisch、Christensen [Christensen et al., 1999] 和 SPES,可快速、可靠、有效地评估肺部病变,数字化工具可简化数据的统计分析。这些评估有助于详细分析病变对胴体重和肉质的影响。宰后肺部评估工具可促进评估过程和后续数据分析。
▶ 1、PRRSV-1是如何引入中国的?
1991年,在荷兰中部城市Lelystad首次发现PRRSV-1,随后命名为Lelystad毒株(LV),该毒株被认为是PRRSV-1的原型株。随后,该毒株传播到法国、比利时、英国、西班牙和德国。迄今为止,尽管PRRSV-1已经传播到加拿大、美国、中国、韩国和泰国,但PRRSV-1依然主要在欧洲流行。
近年来,随着中国养猪产业的发展,中国不断从美国、丹麦、法国、英国、加拿大等地区进口种猪,尤其是丹麦,是我国种猪进口的主要来源之一。而以上所有国家都有PRRSV-1的暴发与流行。由于中国目前禁止使用PRRSV-1疫苗(MLV),因此,近年来PRRSV-1在中国的快速传播,极有可能通过进口种猪的引入导致跨境传播。
已经有研究表明,存在4种来自不同国家的PRRSV-1引入中国。第一,在2006年以前,以类LV为代表的毒株被引入中国并广泛传播,形成了独立的分支。第二,与匈牙利毒株高度相似的PRRSV-1毒株引入中国。第三,类似于美国PRRSV-1疫苗毒株引入中国。第四、在2009年之前,与丹麦PRRSV-1类似的毒株被引进中国。总而言之,PRRSV-1在中国不断被引入和传播。
▶ 2、当前PRRSV-1在中国的发现与流行
早在1997年,中国海关就拦截了感染PRRSV-1(B13,GenBank:AY633973)的猪。这意味着PRRSV-1的引入可能发生在20多年前。直到2011年之前,GenBank中只有少数中国PRRSV-1的部分序列(B13,FJ603)。
2011年,田克恭教授团队从2006-2009年在中国流行的临床-like本中分离出PRRSV-1株(BJEU06-1,NMEU09-1),这是在中国猪场首次报道PRRSV-1野毒。
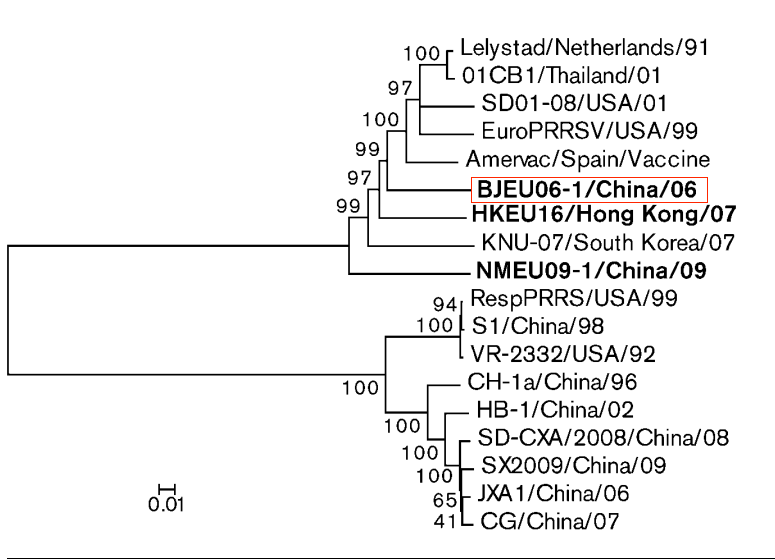
与此同时,2011年,杨汉春教授课题组分离出一株与西班牙疫苗毒株Amervac高度同源的PRRSV-1毒株GZ11-G1。这可能与引进接种过PRRSV-1疫苗的种猪有关。从那时起,PRRSV-1的感染案例在中国不断被发现。特别是近年来,在中国检测到的PRRSV-1毒株数量迅速增加。
2016年,翟少伦从广东省50个猪场采集了750份样本,其中PRRSV-1感染阳性率为24.8%(186/750),这是迄今为止中国PRRSV-1阳性检出率最高的研究报告。此外,从2018-2020年国家/OIE PRRS参考实验室对种猪场和屠宰场检测到PRRSV-1的数据分析发现种猪场检出率高于屠宰场,说明中国PRRSV-1来源仍与引种有关,屠宰场继续检测阳性,说明PRRSV-1已从种猪场扩散至商品猪场。
随着近年来非洲猪瘟在中国的暴发,对PRRSV的监测也得到了加强。这也间接导致了中国PRRSV-1检测报告的增加。截至目前,PRRSV-1感染仍在蔓延,并已在中国至少23个省区流行。PRRSV-1广泛分布于中国的中部、北部、南部、东部、东北和西南部。
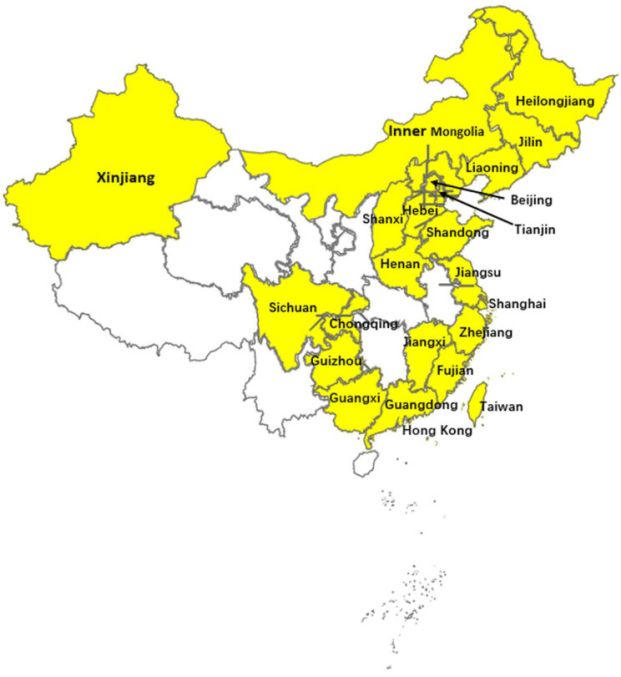
截至2023年3月,中国已经检测到PRRSV-1的区域(黄色)
▶ 3、中国PRRSV-1的分类
分析表明,我国分离的PRRSV-1毒株均属于亚型1。既往研究表明,中国的PRRSV-1可进一步分为4个亚群,包括BJEU06-1-like、Amervac-like、HKEU16-like和NMEU09-1-like(图2)。BJEU06-1-like已成为中国部分地区PRRSV-1的主要亚群。2022年,对河南省一个育肥场进行了PRRSV监测,38份PRRSV阳性-like本中,有14株鉴定为PRRSV-1(36.94%),均属于BJEU06-1- like。对2016-2022年在中国8个省份检测到的20株PRRSV-1毒株的ORF5基因序列进行分析,其中16株PRRSV-1毒株属于BJEU06-1-like。为了探讨中国PRRSV-1各亚组的流行情况,田志军教授团队分析了截至2023年3月上传的中国PRRSV-1 ORF5序列和部分外来PRRSV-1 ORF5参考序列(图3A)。
结果表明,在中国PRRSV-1毒株中,约有38.1%(32/84)属于BJEU06-1-like亚群。过去5年(2018年以来),在中国检测到的PRRSV-1毒株中,BJEU06-1-like毒株占58.1%(18/31)。全基因组遗传演化分析显示,中国PRRSV-1型毒株总数为45.2% (14/31),BJEU06 -1-like毒株占60%(6/10)。这一证据表明BJEU06-1-like毒株是中国最大的PRRSV-1亚群,并已成为中国部分地区PRRSV-1主流毒株。此外,基于ORF5的遗传演化分析,HLJB1和HeB47被发现属于Amervac-like亚群。新的发现还包括,根据ORF5序列的遗传演化分析,有中国PRRSV-1毒株形成3个新的独立亚群。表明中国的PRRSV-1已被发现至少在过去5年中发生了很大的变化。
通过对来自中国和其他国家的PRRSV-1毒株ORF5和全基因组序列的遗传演化分析,结果显示中国的BJEU06 -1-like毒株和NMEUuemn09 -1-like毒株形成了两个截然不同且独立的大亚群,而其他亚群毒株则分散在包括外源PRRSV-1毒株在内的更大亚群中。这些结果表明,BJEU06 -1-like株和NMEU09 -1-like株已在中国稳定存在和传播。
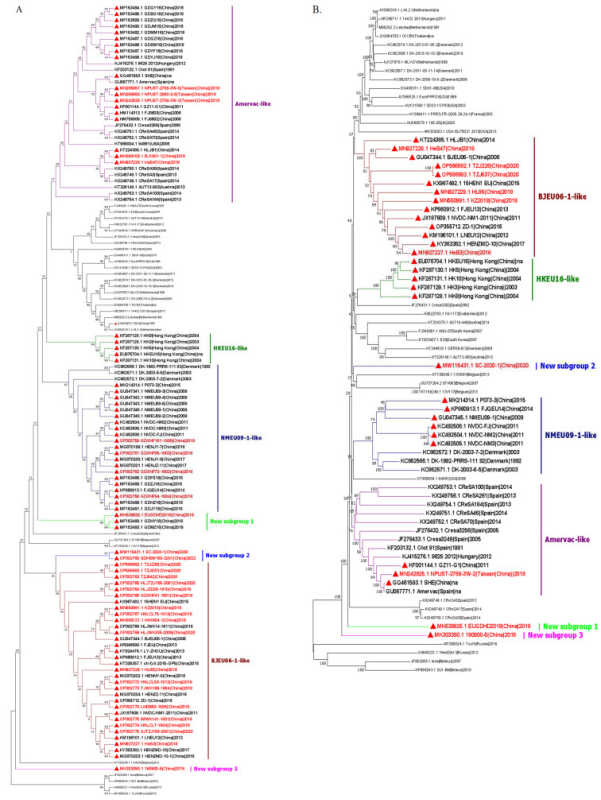
中国PRRSV-1分离株属于亚型1,可分为Amervac-like、BJEU06 -1 like、HKEU16 like和NMEU09 -1 like 4个亚群,并形成3个新亚群。
▶ 4、中国PRRSV-1不同亚群的分布
为进一步明确各亚群在中国的分布特征,利用NCBI已知的来自的中国的PRRSV-1毒株,对不同亚群的分布区域进行了全基因组分析。结果表明,PRRSV-1的主要亚群BJEU06 -1-like亚群传播范围较广,但大多数病例集中在中国北部地区(图3A)。我们推测其原因是中国北方对BJEU06 - 1-like毒株防控不力或中国南方缺乏监测数据。因此,中国各地区应加强对PRRSV-1毒株的预防、控制和监测。目前,HKEU16-like毒株仅在香港发现,尚未传播到中国大陆(图3C)。相比之下,新的独立亚群的毒株分布更为分散(图3E)。
目前发现的亚群以广东省最多。这说明PRRSV-1在广东省的传播较为严重。上述证据表明,PRRSV-1在中国持续传播和突变。因此,应加强海关对进口猪的监测和中国区域内PRRSV-1的监测,阻止PRRSV-1在中国流行
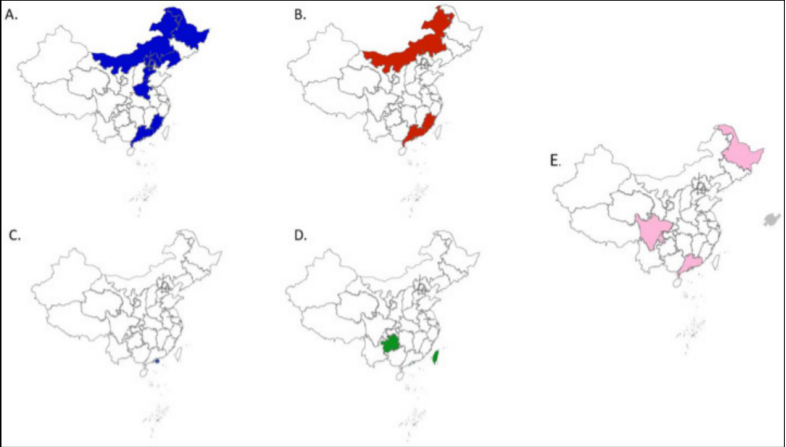
中国PRRSV-1各亚群在的分布
(A)BJEU06-1-like毒株的分布用蓝色标记。
(B)NMEU09-1-like毒株的分布用红色标记。
(C)HKEU16-like毒株的分布用紫色标记。
(D)Amervac-like毒株的分布用绿色标记。
(E)新亚群的毒株分布用粉红色标记。
▶ 5、中国流行的PRRSV-1的基因组特征
PRRSV的遗传变异是RNA病毒中最高的。为了探究PRRSV-1在中国的遗传和进化趋势,作者分析了中国PRRSV-1毒株与已发表的PRRSV全基因组的同源性。结果表明,Nsp2是变异最大的非结构蛋白。在Nsp2区域,中国PRRSV-1株的氨基酸同源性为72.2~99.7%。相比之下,Nsp9和E蛋白分别是最保守的非结构蛋白和结构蛋白。这两个区域的氨基酸差异较小,同源性较高。
为了进一步明确中国PRRSV-1亚型不同亚群之间和各亚群内的变异趋势,对各亚群的核苷酸序列进行同源性分析。结果表明,中国PRRSV-1亚型之间存在较大的变异,基因组同源性为81.5~93.5%。其中,BJEU06 -1-like株和NMEU09 -1-like株具有较大的内部变异,其基因组同源性分别为86.0~99.7%和86.1~99.7%。
另外,构成新独立亚群的3株中国PRRSV-1毒株EUGDHD2018、SC-2020-1、180900-5与其他亚群毒株同源性也较低,同源性为81.5% ~86.8%。而Amervac-like株和HKEU16-like株变异较小,同源性分别为951 - 99.8%和92.6-97.3%。结合地理特征分析上述各亚群变异率较高的亚群,如BJEU06-1-like和NMEU09-1-like更广泛地分布和分散,表明中国的PRRSV-1毒株变异性的增加加剧了PRRSV-1传播风险和防控难度。
▶ 6、中国PRRSV-1的致病性
在以往的案例报道中,与中国PRRSV-2感染病例相比,大多数中国PRRSV-1感染病例的临床症状较轻,致病性较弱,病毒载量较低,临床症状较少。因此,PRRSV-2型在国内得到了更多的关注,而PRRSV-1型的研究一直被忽视。然而,国外已经发现并报道了PRRSV-1亚型1、2和3的高致病性毒株。有学者比较了PRRSV-1毒株BOR59和ILI6亚型2毒株、18794亚型1毒株的致病性。结果表明,BOR59毒株致病性最强,为高致病性毒株,ILI6毒株为中等毒力,18794毒株为低毒力。这说明同一亚型不同毒株的致病性也不同。作为世界上最大的猪肉消费国,中国对种猪及猪肉进口有相当大的需求,这表明高致病性PRRSV-1可能有通过贸易传入中国的风险,类似于过去非洲猪瘟的传入方式。
虽然中国分离的PRRSV-1毒株报告数量逐渐增加,但对中国PRRSV-1株的病原学鉴定仍然有限。据我们所知,中国关于PRRSV-1致病性的报道仅有6篇:HLJB1、GZ11-G1和NPUST 2789-3W-2属于Amervac-like亚群,ZD-1和SD1291属于BJEU06 -1-like亚群,181187-2与PR40/2014形成一个新的独立亚群。6株毒株均表现为中、低致病性。GZ11-G1毒株对仔猪不致命,但具有致病性,可引起仔猪呼吸道症状和短暂性体温升高。它在体内的复制效率高于Amervac毒株。HLJB1在感染猪体内保持低病毒血症并诱导良好的免疫应答。NPUST 2789-3W-2毒株具有较低的致病性,高毒量接种组和接触组实验仔猪血清病毒载量分析表明,台湾野生型PRRSV-1具有潜在的高传染性。181187-2与意大利分离的高致病性毒株PR40/2014属同一亚型,具有中等致病性。
作者推测它可能是在引进种猪时从意大利传入。SD1291毒株是一个致病性相对较低的分离株,但其体外复制效率明显高于从PAMs中分离的HLJB1毒株。ZD-1毒株对仔猪具有中等致病性,在GP3的c端有26-aa的提前截断。高致病性PRRSV-1毒株Lena在GP3的c端也有过早终止。然而,目前尚不清楚GP3的这些过早终止是否对致病性有任何影响。
比较不同PPRRSV-1毒株的致病性数据,目前中国流行的PRRSV-1毒株的致病性总体较低,但2023年报道的181187-2毒株和ZD-1毒株的出现表明中国PRRSV-1的致病性有轻微上升的趋势。此外,已发现一株高致病性PRRSV-1毒株在邻国和重要生猪进口国均有报告,我们应加强对PRRSV-1毒株的监测,并设法避免从高致病性PRRSV-1暴发国家进口生猪。
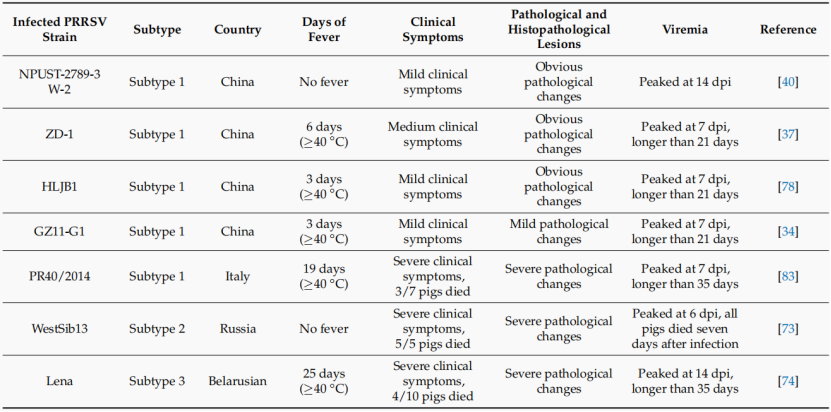
不同PRRSV-1的致病性比较
▶ 7、PRRSV-1的诊断方法
PRRSV检测在临床预防和控制中具有重要作用。除了通过直观的临床症状进行诊断外,目前还有一系列基于核酸和抗原/抗体的检测方法可用于PRRSV的检测,包括病毒分离(VI)、间接免疫荧光法(IFA)、单层免疫过氧化物酶法(IPMA)、血清病毒中和试验(SVN)、反转录PCR (RT-PCR)、实时荧光定量PCR (qPCR)、数字PCR (dPCR)、重组聚合酶扩增(RPA)、环介导等温扩增(LAMP)、CRISPR、新一代测序(mNGS)和酶联免疫吸附测定(ELISA)。随着分子生物学和基因测序技术的快速发展,RT-PCR检测方法逐渐成为PRRSV抗原检测方法的主流。同时,开发了巢式PCR、多重PCR、实时荧光定量PCR等检测方法。目前国内已有建立多种RT-PCR检测方法区分PRRSV-2和PRRSV-1的报道。这些检测方法耗时短,速度快,适用于急性发病猪PRRSV的实验室诊断。然而,当遇到变异较大的PRRSV毒株时,这些方法容易出现误检或漏检。
目前,ELISA具有简便、快速、灵敏、易于标准化等优点,广泛应用于PRRSV血清抗体的检测。特别适合进行大规模流行病学调查和监测。目前市面上大量的ELISA试剂盒,大多采用PRRSV N蛋白(Jinnuo和IDEXX)或膜蛋白(Hipra)作为间接ELISA的包被抗原。IDEXX PRRSV检测方法是世界范围内最常用的方法,被普遍认为是PRRSV抗体检测的金标准。然而,这些常用的商用ELISA试剂盒并不提供PRRSV-1和PRRSV-2特定性检测。Hipra声称他们的两种ELISA方法可以检测PRRSV-1和PRRSV-2抗体。然而,有报告显示缺乏特异性。
另一方面,Venteo等人开发了一种基于PRRSV-1核衣壳蛋白ELISA方法,该蛋白已显示出检测PRRSV-1早期感染的能力。目前,PRRSV的诊断方法在中国主要侧重于PRRSV-2感染的检测,而目前现有的PRRSV-1感染检测方法存在局限性。这种差异表明有必要进一步发展和改进专门针对PRRSV-1的诊断技术。通过扩大可用检测方法的范围,准确识别和鉴定的能力可以改善中国PRRSV-1感染的监测,从而更好地控制和预防PRRSV-1感染。
▶ 8、PRRSV-1的预防控制
PRRSV-1疫苗研发
由于PRRSV的高变异性和传染性及在世界范围内广泛流行。目前,用于预防和控制PRRS的主要商业化或正在开发的疫苗包括灭活疫苗、MLV、亚单位疫苗、DNA疫苗和病毒载体疫苗。其中,世界上最常用的PRRSV疫苗是PRRSV MLV,是1994年在美国推出的第一种商业化PRRSV疫苗。在20世纪90年代末,PRRSV-1 MLV在欧洲被用于预防PRRSV-1感染。
到目前为止,它在一些亚洲地区,如越南、韩国和中国台湾也被允许。全球常见的PRRSV-1 MLV包括Porcilis PRRS (MSD Animal Health)、Amervac PRRS (Hipra)、ReproCyc PRRS EU(BI)、Pyrsvac-183 (SYVA Laboratories)、Ingelvac PRRSFLEX®E(BI)、Suvaxyn®PRRS MLV (Zoetis)和Unistrain®PRRS (Hipra)(表5)。PRRSV MLV可以减少病毒在感染猪中的传播,并诱导对同源野生型毒株有效的保护性免疫反应。
然而,PRRSV MLV对异源毒株仅提供部分保护或没有保护。此外,PRRSV MLV在复制过程中有产生疫苗毒株或与野生毒株重组的风险。因此,这些疫苗的安全性受到了挑战。疫苗重组毒株的出现无疑增加了中国预防和控制PRRSV-1感染的难度。因此提醒我们,合理使用疫苗,进一步加强国内对PRRSV-1 MLV的管理和输入性动物疾病的监测是非常重要的。
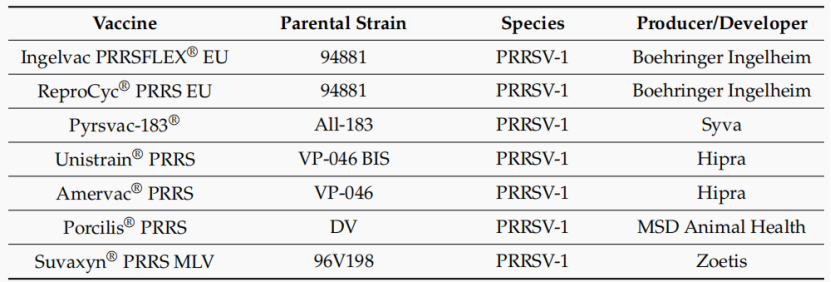
国外主要使用的PRRSV-1商品化疫苗
PRRSV-1的预防与控制
随着中国地区PRRSV-1遗传多样性和变异多样性的增加,新毒株出现频繁,现有毒株亚群复杂,加大了中国PRRSV-1防控的难度。由于PRRSV-1在中国流行的致病性较低,因此受到的关注较少。因为缺乏有效的临床对PRRSV-1的检测方法不完善,可能导致PRRSV-1漏检。
目前,中国预防和控制PRRSV感染的重点是通过疫苗接种预防和控制PRRSV-2感染。然而,中国针对PRRSV-1的诊断试剂和疫苗储备不足,预防和控制PRRSV-1感染主要采用生物安全管理、建立良好的饲养机制,营养均衡、保持猪场适宜的饲养环境和确保合理的饲养密度。以及有必要严格控制生猪流动,确保生猪运输作业标准化等措施。另外,各地区加强PRRSV-1定期监测,以及需要更多的研究来更好地了解中国PRRSV-1来实现有效的防控。(作者:李智 博维特)






